La respirazione cellulare si divide in tre fasi principali
- Ossidazione di molecole organiche in Acetil-CoA
- Ciclo di Krebs, o dell’acido citrico
- Fosforilazione ossidativa
Tutte e tre avvengono nei mitocondri in cui sono trasportati tramite trasporto attivo secondario. In questo articolo ci occuperemo del ciclo di Krebs.
Cos’è il ciclo di Krebs
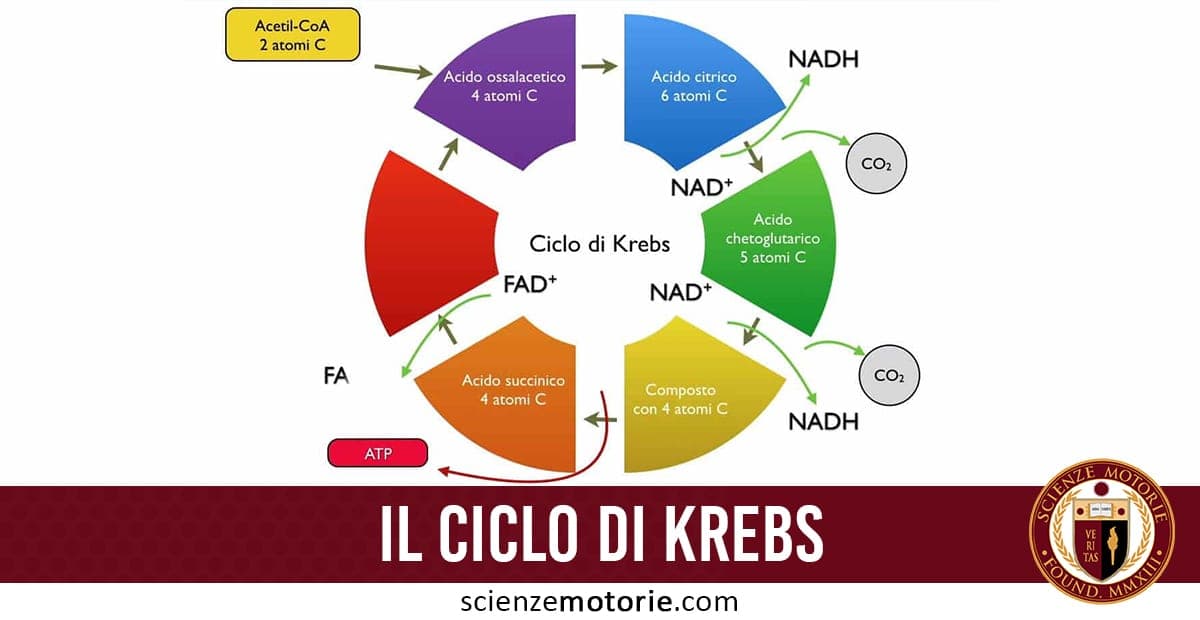
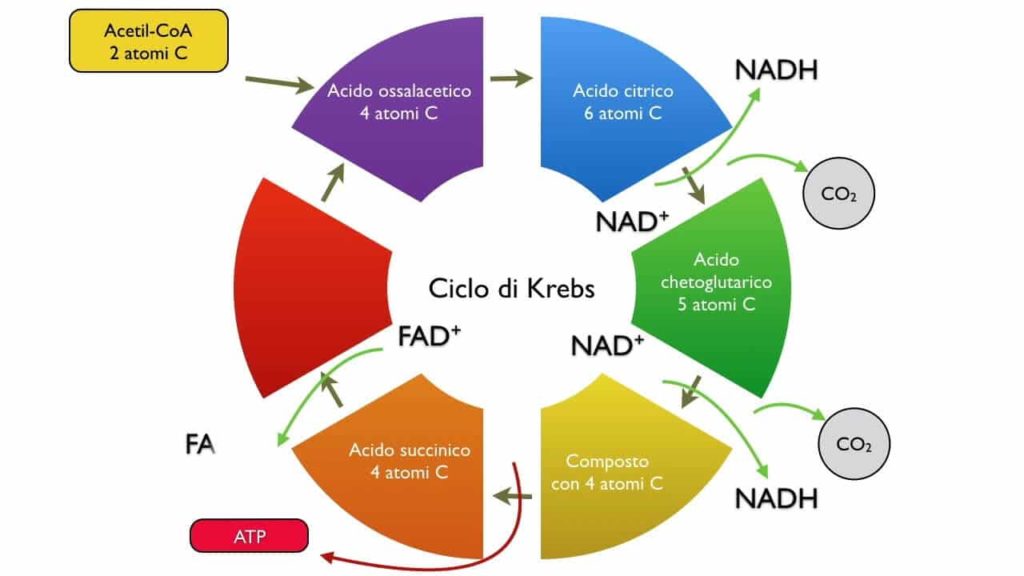
Il ciclo dell’acido citrico è anche noto come ciclo di Krebs o ciclo dell’acido tricarbossilico ed è caratterizzato da 8 tappe, anch’esso non è prettamente energetico, ma prepara i trasportatori di elettroni alla fosforilazione ossidativa.
È una serie di reazioni in un circuito chiuso che sono fondamentali per la respirazione cellulare. Il ciclo dell’acido citrico produce la molecola ad alta energia ATP (adenosina trifosfato) e i sottoprodotti che formano anche l’ATP in un ulteriore processo chiamato fosforilazione ossidativa.
La regolazione del ciclo dell’acido citrico è importante in quanto le reazioni che non sono controllate porteranno a grandi quantità di energia metabolica sprecata. La capacità di regolare il ciclo mantiene la cellula in uno stato stabile e questa funzione è mantenuta da tre meccanismi:
- La disponibilità di substrati.
- Inibizione dei prodotti formati
- Inibizione degli enzimi attraverso il feedback allosterico.
Regolazione dell’acetile CoA
Molecole organiche dotate di energia (carboidrati, lipidi, ma anche proteine), vengono scisse in precedenti reazioni e, prima di entrare nel ciclo di Krebs, vengono trasformate in acetilCoA, una molecola formata da un gruppo acetile (CH3CO-) e da un trasportatore di acili, detto coenzima A.
Nella prima fase del ciclo di Krebs, l’acetil-CoA viene legato ad un composto a 4 atomi di carbonio, l’ossalacetato, che quindi forma un composto a 6 atomi di carbonio, il citrato (da cui il nome alternativo del ciclo). Quest’ultimo viene avviato poi in una serie di reazioni che consentono la sintesi di 3 molecole di NADH, 2 molecole di FADH2 ed una molecola di ATP, derivante dalla defosforilazione del GTP. Per ossidare una molecola di glucosio sono necessari quindi 2 giri del ciclo di Krebs.
Il flusso del metabolita è inibito allostericamente, dove un enzima è regolato legando una molecola effettrice ad un sito non attivo. La reazione del complesso piruvato deidrogenasi è inibita allostericamente quando vi sono alti rapporti di ATP in ADP, NADH in NAD + e acetil-CoA in CoA. L’attivazione allosterica si verifica quando i volumi del rapporto diminuiscono.
Regolazione degli enzimi nel ciclo dell’acido citrico
Tre reazioni del ciclo sono catalizzate rispettivamente dagli enzimi:
- Citrato sintasi.
- Isocitrato deidrogenasi.
- α-chetoglutarato deidrogenasi
Prima fase
La citrato sintasi è responsabile della velocità di reazione nella prima fase del ciclo quando l’acetil-CoA è combinato con acido ossalacetico per formare citrato. È inibito da alte concentrazioni di ATP, acetil-CoA e NADH che indicano un già alto livello di approvvigionamento energetico. La molecola prodotta nella reazione, il citrato, può anche agire da inibitore della reazione.
Poiché la citrato sintasi è inibita dal prodotto finale del ciclo dell’acido citrico come ATP, l’ADP (adenosina difosfato) agisce come attivatore allosterico dell’enzima quando l’ATP è formato da ADP. Pertanto, la velocità del ciclo si riduce quando la cella ha un alto livello di ATP.
Seconda fase
Nella seconda fase il citrato viene isomerizzato ad isocitrato per poi essere decarbossilato ad a-chetoglutarato (a-KTG). Avviene prima una deidratazione seguita da una idratazione. La reazione viene catalizzata dall’enzima aconitasi, enzima complesso contenente ferro e zolfo nel centro attivo.
Il citrato viene convertito in isocitrato attraverso la formazione dell’intermedio cis-aconitato. L’enzima citrato sintasi viene inibito allostericamente dall’ATP.
Terza fase
L’enzima isocitrato deidrogenasi è un catalizzatore importante nella terza fase della reazione. Regola la velocità alla quale l’isocitrato isomero citrato perde un carbonio per formare la molecola a cinque carbonio α-chetoglutarato. Il coenzima NADH è un prodotto della reazione e, a livelli elevati, agisce come un inibitore spostando direttamente le molecole NAD + da cui è formato.
Quarta fase
L’enzima α-chetoglutarato deidrogenasi è un altro importante catalizzatore nella quarta fase del ciclo in cui anche l’α-chetoglutarato perde un carbonio e si combina con il coenzima A per formare succinil CoA. I due prodotti della reazione, succinil CoA e NADH, funzionano entrambi come inibitori a grandi concentrazioni.
Quinta fase
La quinta fase è catalizzata dalla succinil-CoA sintetasi ed è molto importante perché è l’unica tappa in cui viene prodotto un nucleotide diverso dall’ATP. Tale enzima separa il CoA dal succinil-CoA sotituendolo con un fosfato libero. Verrà quindi rilasciato il succinato e il gruppo fosfato legherà un GDP per formare una molecola di GTP.
Sesta fase
Nella sesta fase il succinato viene ossidato nei carboni 2 e 3 formando un doppio legame tra i due con la formazione di un FADH2. L’enzima è la succinato deidrogenasi e la molecola formata è il fumarato.
Settima fase
Nella settima fase Il doppio legame viene idratato tramite la fumarasi formando L-malato. L’enzima è altamente stereospecifico e l’isomero che può formarsi è solo il “trans L”.
Ottava fase
Nell’ottava fase il gruppo ossidrilico viene ossidato dalla malato deidrogenasi formando nuovamente ossalacetato e un NADH.
Calcio come regolatore del ciclo dell’acido citrico
Il calcio è anche un importante regolatore del ciclo di Krebs; un aumento delle concentrazioni sia di ADP che di ioni di calcio (Ca 2+) sono una conseguenza dei cambiamenti nell’attività cellulare. Pertanto, il segnale che stimola la contrazione muscolare attiva anche la produzione dell’ATP che lo sostiene, attraverso il ciclo dell’acido citrico. Gli ioni calcio regolano il ciclo dell’acido citrico attivando la piruvato deidrogenasi, il primo componente della reazione complessa della piruvato deidrogenasi che forma acetil-CoA.
Gli ioni calcio attivano anche gli enzimi, l’isocitrato deidrogenasi e l’α-chetoglutarato deidrogenasi che catalizzano rispettivamente la terza e la quarta fase del ciclo. L’attivazione di questi enzimi, attraverso gli ioni di calcio, aumenta il tasso di reazioni separate all’interno del ciclo e quindi aumenta la produzione del prodotto per l’intero ciclo.
Riferimenti:
- Berg JM, Tymoczko JL, Stryer L. Biochemistry. 5th edition. New York: W H Freeman; 2002. Chapter 17, The Citric Acid Cycle. https://www.ncbi.nlm.nih.gov/books/NBK21163/
- Berg JM, Tymoczko JL, Stryer L. Biochemistry. 5th edition. New York: W H Freeman; 2002. Section 17.2, Entry to the Citric Acid Cycle and Metabolism Through It Are Controlled. https://www.ncbi.nlm.nih.gov/books/NBK22347/
- Wan, B. et al. 1989. Regulation of citric acid cycle by calcium, Journal of Biological Chemistry, 264, pp. 13430-13439. https://www.ncbi.nlm.nih.gov/pubmed/2503501
- Traaseth, N. et al. 2004. Role of calcium signaling in the activation of mitochondrial nitric oxide synthase and citric acid cycle, Biochimica et Biophysica Acta (BBA) – Bioenergetics, 1658, pp.64-71. http://www.sciencedirect.com/science/article/pii/S0005272804001343













